Innate Immunity in the Randow Lab







We are interested in cell autonomous innate immunity, i.e. in the ability of individual cells to defend themselves against infection.
Cytosolic defense against bacterial invasion
The cytosol is rich in nutrients but very few bacteria can colonize it. We wish to understand how cells employ the ubiquitin system and autophagy to defend their cytosol.
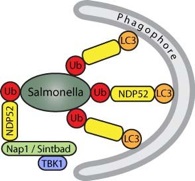
Salmonella entering the mammalian cytosol can become coated with ubiquitin and we have identified the cellular receptor for such ubiquitin coated bacteria, a protein called NDP52. Upon binding the bacterial ubiquitin coat, NDP52 recruits the kinase TBK1 and, by binding to ATG8/LC3, delivers the bacteria into autophagosomes for destruction. NDP52 is, in fact, a novel autophagy receptor.
Regulation of NF-kB and IRF signaling
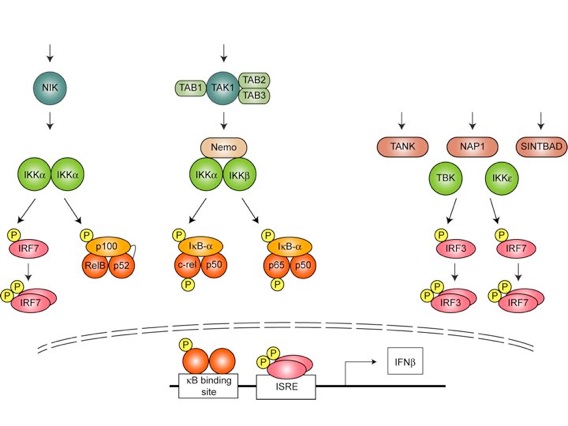
The defense against pathogens also requires profound changes in gene expression, which are coordinated by latent transcription factors of the NF-kB and IRF families. We wish to understand in mechanistic detail how infection activates NF-κB and IRF signaling.
The genetic analysis of somatic cell lines holds significant potential to our understanding of signal transduction (and other cell autonomous traits). Through homologous recombination and chemical mutagenesis we have obtained a large collection of gene deficient human and murine cells. Using these cells we focus on the very upstream and the very downstream components of the pathways – how do Toll-like receptors acquire signaling competence and how do adaptor proteins and ubiquitin control the activity of IKK family kinases?
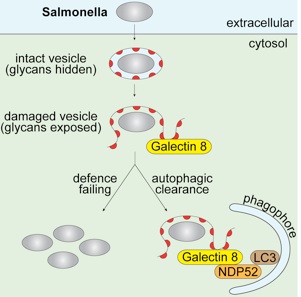
More recently we identified a ubiquitin-independent function of NDP52. In this new pathway NDP52 binds to Galectin 8, a cytosolic lectin that detects host glycans on vesicles damaged by Salmonella in the process of entering its host cell. Galectin-8 senses damage to host vesicles even under sterile conditions (click here to see a movie). Galectin-8 is therefore a novel danger receptor that monitors the integrity of the endosomal / lysosomal compartment and detects a wide variety of vesicle-damaging pathogens in addition to Salmonella, for example Shigella and Listeria.
We now also understand why NDP52 binds exclusively to Galectin-8; sterical hindrance prevents its binding to most other galectins (click here to listen