BAR-Superfamily
|
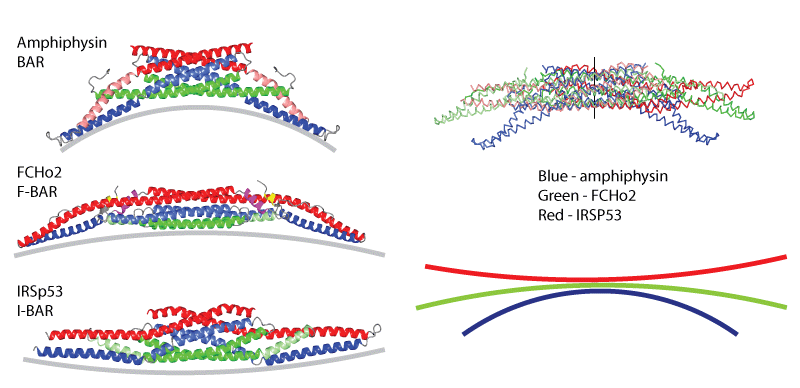
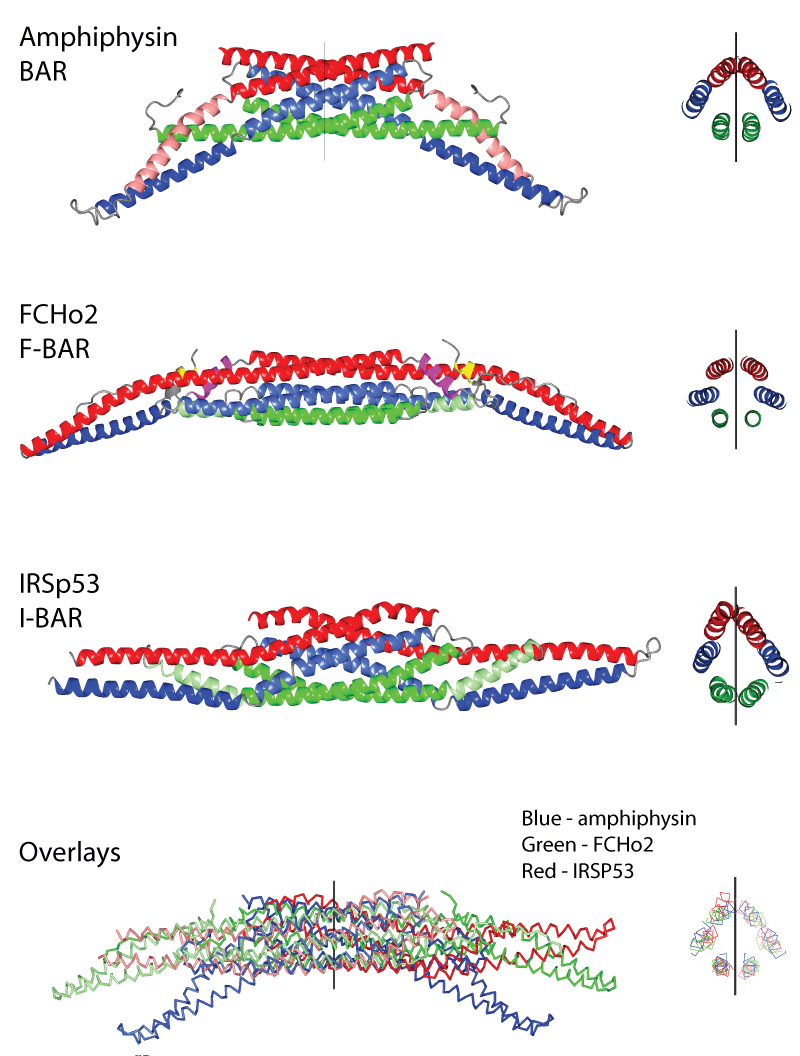
What makes up the BAR Superfamily? These are proteins that bind to membrane with different curvature preferences. The BAR superfamily can be subdivided into BAR/N-BAR modules that bind to membranes of high positive curvature, F-BAR modules that bind to a different range of positive membrane curvatures and I-BAR modules that binding to negatively curved membranes (see curvature profiles below). |
 |
| See other views of Superfamily members |
A brief History....
BAR
-
BAR recognised as a major N-terminal region of homology between amphiphysin relatives (BIN1, Amphiphysin and RVS167) (Sakamuro et al Nat Genetics Sept 1996)
-
Amphiphysin can tubulate liposomes in vitro like dynamin; implications for membrane bending (Takei et al. Nat Cell Biol; May 1999)
-
First BAR module structure (Amphiphysin BAR) and characterization of the “BAR protein family” (Peter et al. Science; Jan 2004). Recognition from this structure that BAR modules are found in many different proteins and recognition of a subgroup of BARs with an N-terminal amphipathic helix, which we called N-BAR domains. We also recognised that a previous structure of arfaptin could now be reclassified as a BAR domain (Tarricone et al. Nature; May 2001)
-
Endophilin-A1 BAR domain structure (Weissenhorn JMB; Aug 2005)
-
Endophilin mechanism of membrane bending and 2nd membrane-inserting helix analysis (Gallop et al. EMBO J; Jun 2006 & Masuda et al. EMBO J; Jun 2006)
-
BAR-PH structure of APPL1 and biochemical analysis (Li, et al. Structure. May 2007) & (Zhu et al. EMBO J; Jul 2007)
-
PX-BAR structure of sorting nexin 9 presented with biochemical and in vivo analysis (Pylyenko et al. EMBO J; Oct. 2007)
|
F-BAR
-
“FCH domain” first recorded as a region of homology in FER and CIP4 proteins, and attributed to a possible actin interaction domain (Aspenstrom. Curr Biol; Jul 1997)
-
FBP17 tubulates in vivo and can dimerize via an “extended FCH” (EFC) region (Kamiota et al. JBC; Jul. 2004)
-
In vitro tubulation and PS/PI lipid preference of several “F-BAR” domains, and further analysis of their interplay with actin/dynamin machinery (Itoh, et al. Dev Cell; Dec 2005)
-
Characterized in vitro/in vivo tubuation of several EFC/F-BAR domains, including FBP17, CIP4, PSTPIP1/2, and FER (Tsujita et al. JCB; Jan 2006)
-
Syndapin dimers/oligomers contain BAR-like domain and interplay with vesicle formation (Kessels et al. JBC; May 2006)
-
Toca1 in vivo tubulation via F-BAR region and evidence for membrane trafficking in hippocampal neurons (Kamimoto et al. JBC; Sept. 2006)
-
FBP17 & CIP4 F-BAR domain structures and proposed in vivo function as vesicle sculptors proposed (Shimada, et al. Cell; May 2007)
-
FCHo2 F-BAR domain structure and biochemical analysis (Henne et al. Structure; May 2007)
|
I-BAR
-
IRSp53 and Mena form complex to bend membrane into filopodia (Krugmann et al. Curr Biol; Oct 2001)
-
Initial biochemical analysis of IMD/I-BAR region as an actin bundler (Yamagishi et al. JBC; Jan 2004)
-
IMD/I-BAR structure of IRSp53; proposed actin bundling function (Millard et al. EMBO J; Jan 2005)
-
MIM protein IMD/I-BAR domain domain membrane binding demonstrated and generation of positive opposite curvature suggested (Lee et al. Structure; Feb 2007)
-
I-BARs of IRSp53 & MIM structures analyzed and I-BAR membrane bending mechanism demonstrated via electron cryo-tomography (Mattila et al. JBC; Mar 2007)
|
See F-BAR modules
See I-BAR modules
|

{kind=link}