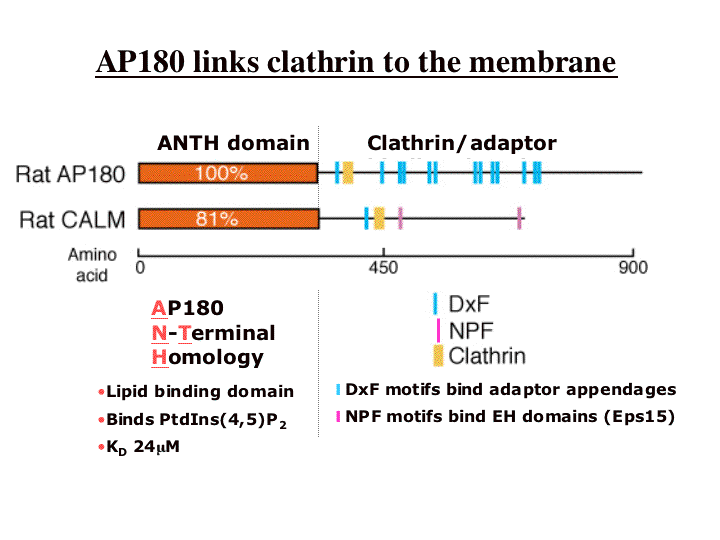
|
AP180 is a brain enriched protein essential for the recycling of synaptic vesicles after exocytosis and CALM is a ubiquitous homologue. AP180 co-purifies with clathrin-coated vesicle (see purification) and we have shown that it functions to tether clathrin to phosphatidylinositol(4,5)bisphosphate (PtdIns(4,5)P2) containing membranes. Thus in the domain structure above there is a membrane binding domain (ANTH domain) and a clathrin/adaptor binding domain. The adaptor binding is discussed further in other parts of these Web pages but it is not as strong as the clathrin binding, and the dominant phenotype of overexpression is a disruption of clathrin-assembly. Of all clathrin-binding proteins we have tested, AP180 is the most efficient at recruiting clathrin to membranes.
The stars (*) in the picture above point to the fact that all the clathrin is recruited to the membrane and in experiments where we titrate in more clathrin then it continues to be all recruited until there is a 1:1 stoichiometry between AP180 and clathrin on the membrane. This is because AP180 promotes the polymerisation of clathrin on the membrane. We visualise this by electron microscopy below:
In COS cells, overexpression of the C-terminus of AP180 (lipid binding domain is missing) leads to complete inhibition o f transferrin uptake. As clathrin-polymerisation is so important in budding from endosomes and from the TGN, this construct can be used as a general inhibitor for clathrin pathways (browse through our experiments ). In these Web pages we see the disruption of AP1, AP2, EEA1, mannose-6-phosphate receptor and TGN46. Note the complete loss of the M6P receptor implies that there is an upregulation or redirecting of pathways to the lysosome. Thus AP180 C-terminus is 'a real killer' for clathrin budding pathways. |