|
|
Epsin1 ENTH domain
|
|
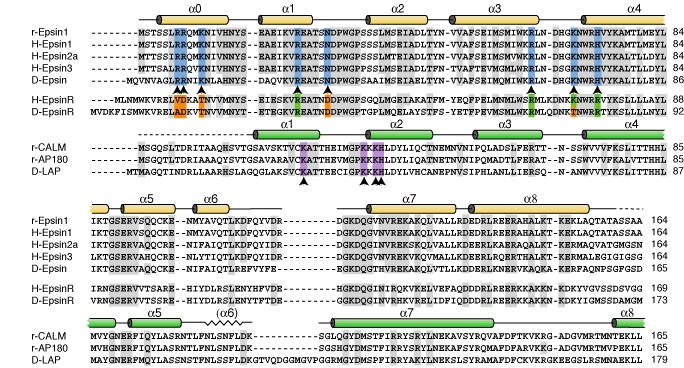
This is the conserved N-terminal domain which in epsin1, 2 and 3 binds PtdIns(4,5)P2. The sequence homology is shown below with the key PtdIns(4,5)P2 binding residues (0). This compares with the PtdIns(4,5)P2 binding resides in the ANTH (0) domain of AP180/CALM and the PtdInsP binding residues in epsinR (0)
|
 |
 |
| These are the key PtdIns(4,5)P2 binding residues of ANTH domains compared with the same region in ENTH domains. These regions act as 'signatures' of ENTH and ANTH domains in proteins. |
|
|
|
|
|
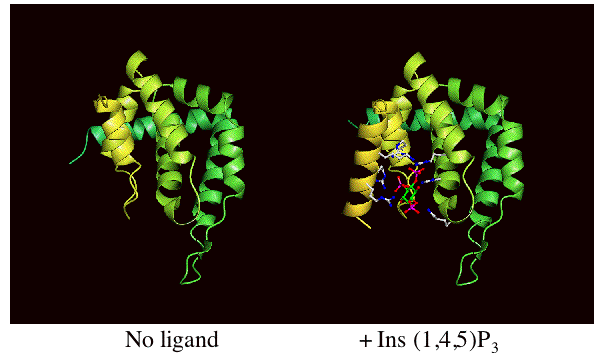
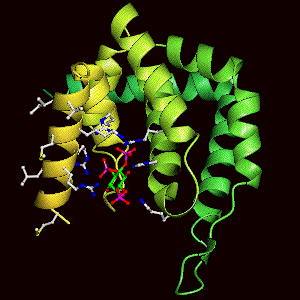
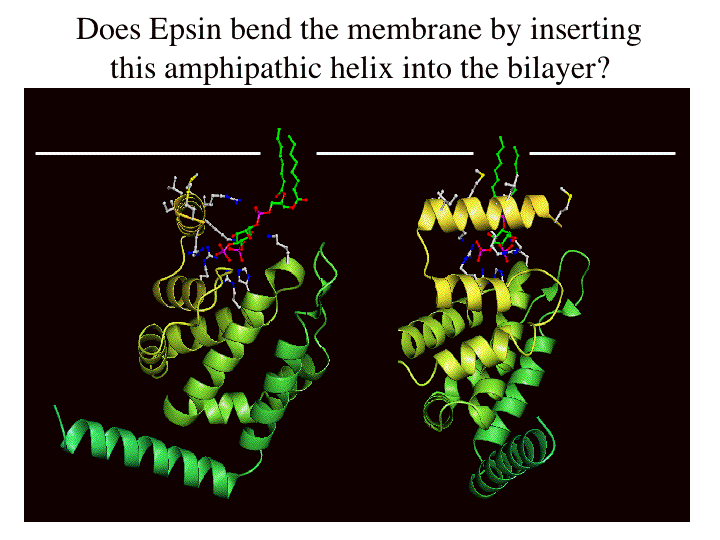
Binding of the lipid headgroup causes the de novo folding of helix zero around the headgroup. This helix folds in vitro, not only on headgroup binding, but on membrane interaction to form the PtdIns(4,5)P2 binding pocket. The folding can be followed by CD spectroscopy. See also other structure pictures.
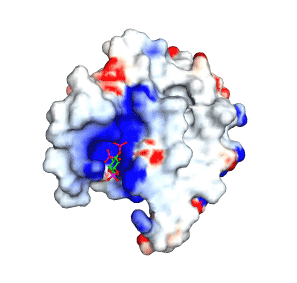
Epsin1 prefers PtdIns(4,5)P2 containing membranes (see coordination and see affinities of headgroups) and
thus even the ENTH domain also localises to domains on the plasma membrane (see immunofluorescence).
|
|
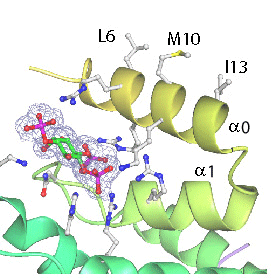
Helix Zero has a hydrophobic outer surface (see below) which we propose interacts with the hydrophobic phase of the lipid bilayer.
|
|
|
Epsin1 tubulates liposomes and drives the invagination of clathrin-coated pits (see EM images).
Thus:
|
|
|
|
|
|
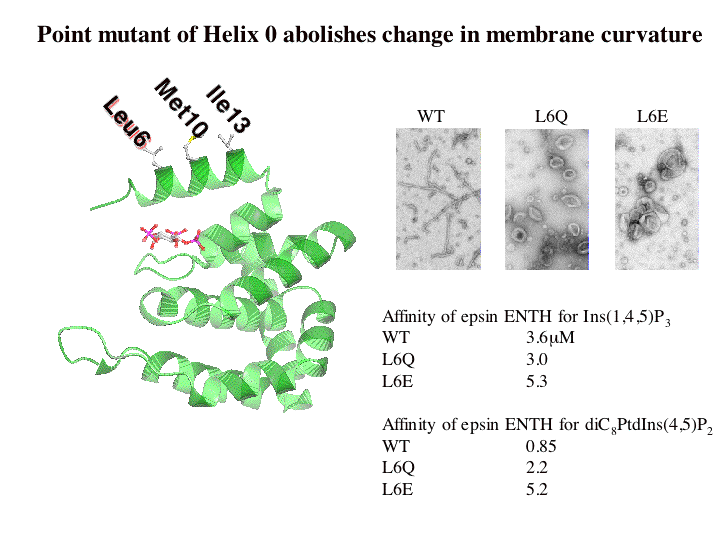
Given that mutations of Leucine6 to more charged residues (L6E and L6Q) prevent membrane insertion of helix zero while the L6W mutant leads to better insertion and liposome vesiculation, we conclude that helix zero is indeed responsible for the membrane bending consequences of epsin binding.
|
|
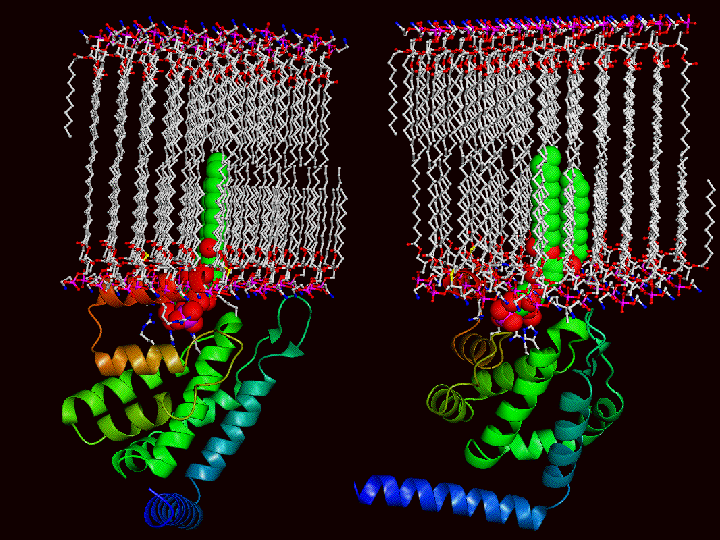
The Model of how the ENTH domain works:
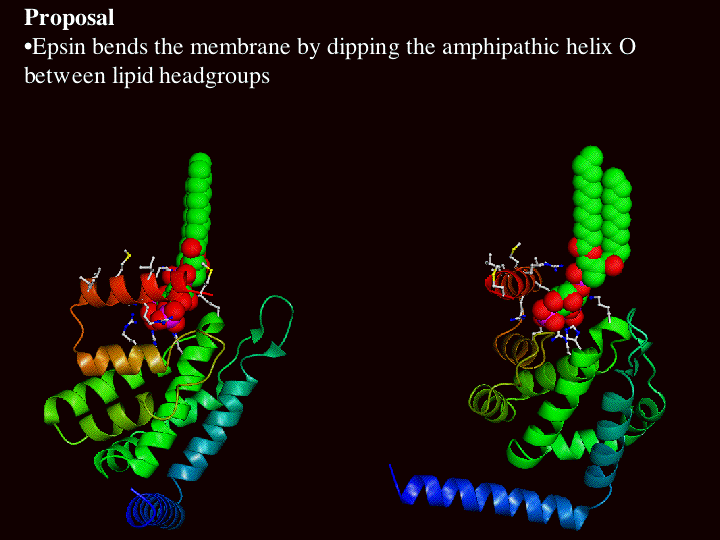
The following pictures illustrate how the lipids must move apart to accommodate the insertion of helix zero. The first picture shows 2 different views of the ENTH domain with a space filled PtdIns(4,5)P2 docked in the structure. In the bottom picture a membrane (to scale) is added such that the PtdIns(4,5)P2 is inserted and consequently helix zero is buried. One can image a space filled helix zero an
d the number of lipids that would have to be displaced to accommodate epsin docking on the membrane
|
 |
 |
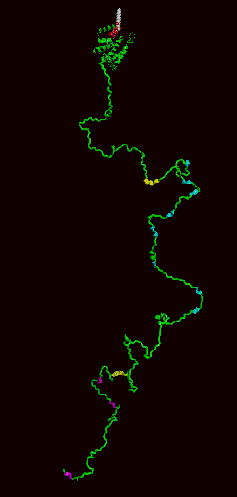 |
Model of epsin1 full-length protein:
The ENTH domain is the alpha-helical domain at the top and the rest of the protein is represented as unstructured and containing multiple clathrin, adaptor and Eps15 binding motifs. The unstructured nature is likely an overexageration as some of our affinity measurements show that binding motifs that are not close to each other in this representation can influence each others binding characteristics.
|
|
| The unliganded ENTH domain was previously solved by Hyman et al 2001 and proposed to interact with a transcription factor. In the same issue of Science as our own paper there was also an NMR study of epsin ENTH domain proposing that it
bound to PtdIns(4,5)P2 (Itoh et al 2001). |
Our paper: Ford, M.G.J., Mills, I.G., Peter, B.J., Vallis, Y., Praefcke, G.J.K., Evans, P.R. and McMahon, H.T. (2002) Curvature of clathrin-coated pits driven by epsin. Nature 419: 361-366 (abstract). (.pdf) supplemental data
(see also News and Views and highlights in Nature Cell Biology, Nature reviews in molecular cell biology, and a mini review in Cell (Cell October 18, 2002: 111 (2):143-146)(
summary)
|
|
|
|
|
|