|
Amphiphysin
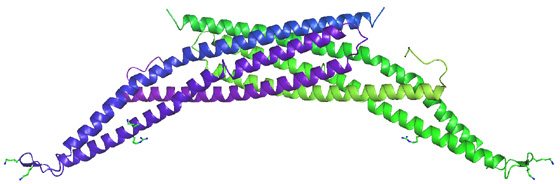
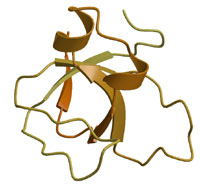
Amphiphysin was first implicated in the recruitment of dynamin by de Camilli and colleagues. Evidence later accumulated that amphiphysin may indeed be the recruitment protein for dynamin to sites of clathrin-mediated endocytosis. Despite the low affinity of the amphiphysin-SH3 domain for dynamin (Owen et al 1998 EMBO J.) the interaction is likely to be considerably stronger in vivo where amphiphysin is a dimer and dynamin is an oligomer (see Wigge et al 1997 MBC). What makes amphiphysin a good candidate protein for dynamin recruitment? In brain extract amphiphysin is found in a 1:1 stoichiometry with dynamin and this interaction is inhibited by phosphorylation and both proteins are dephosphorylated on stimulation of exo-endocytosis. The amphiphysin SH3 domain when overexpressed in COS7 cells is a potent inhibitor of dynamin action when compared alongside other SH3 domains which binds to dynamin (Grb2, Spectrin and PLCg) (see Wigge et al 1997 Current Biol.). The SH3 domain also blocks endocytosis in the Lamprey synapse (Shupliakov et al 1997) leading to the accumulation of endocytic profiles with 'necks' along the plasma membrane. The potency of the isolated SH3 domain is likely due to its ability to disassembly dynamin multimers (
Owen et al 1989 EMBO J.). The binding site for amphiphysin has been carefully mapped to a sequence ProSerArgProAsnArg in the PRD of dynamin. Peptides that specifically inhibit the dynamin-amphiphysin interaction inhibit clathrin-mediated endocytosis. A mutation in a critical lipid binding residue of the PH domain (Lys535Ala) of dynamin leads to a dominant negative phenotype in cells (see Vallis et al 1999) but this can be partially rescued by a second mutation of the amphiphysin binding sequence in the PRD (see Vallis et al 1999) thus showing that the protein that interacts with this sequence plays an important role. |
{kind=link}