Endocytic Interactome Home Page |
Endocytic Interactome:
-
The strength of network biology is its ability to derive information about biology without a priori mechanistic or molecular knowledge.
However a careful understanding of biology can refine the strengths of the network approach.
We do this by taking a well-studied biological pathway, clathrin-mediated endocytosis (CME), and show how the protein machinery is organised around hubs resulting in a pathway interactome where the hub proteins are recognised by their connectivity and their higher conservation over accessory proteins. This approach causes us to observe the design principles behind the pathway and we propose that the principles we derive may be applied to other biological systems. Some of the ideas in this page were first presented at a Gorden Conference in 2006 (see talk) and these pages are to accompany an Analysis written for Nature in 2007 (Main.pdf, Sup.pdf).
|
CME Interactome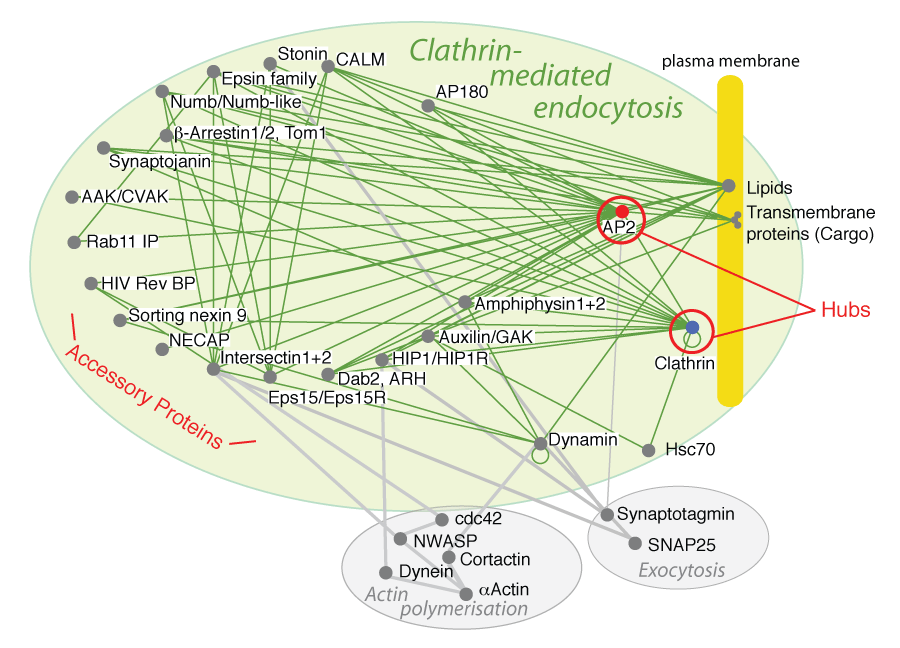
The interactions of proteins involved in clathrin-mediated endocytosis can be plotted, and we get a static view of the complexity of this Pathway Protein Interactome, PPI (see other endocytosis.org pages for descriptions of proteins). The same types of PPIs can be drawn for any pathway where most of the protein components are known.
|
Hubs and nodes:
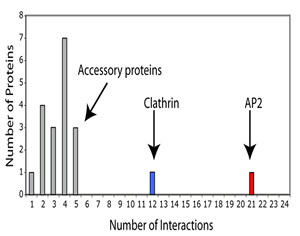
Nodes are the protein interaction partners in the network and hubs are protein nodes with disproportionately large numbers of interactions. In the endocytosis interactome these proteins are clathrin and AP2 adaptors.
Hubs have been subdivided into party- and date- hubs (Han et al 2004, Nature 430). Date-hubs are biomolecules that have many partners but interact with them at different times or locations (dynamic-hubs), while party-hubs can interact with many biomolecules at once (static-hubs). Date-hubs are proposed to organise the proteome, connecting biological processes to each other, whereas party-hubs are proposed to function within modules (in our case, pathways). While recognising the descriptive benefits of ‘party-hub’ and ‘date-hub’ nomenclature we call clathrin and AP2 ‘pathway-hubs’ as this does not immediately ascribe any behaviour to these hubs, but simply states that they appear within the studied pathway. Notwithstanding we do make an observation on the maturation of hubs in our pathway:
|
 |
Hubs formed by clustering proteins
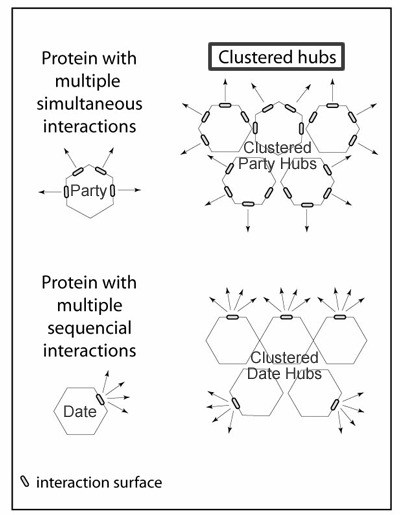 |
Clustered-hubs are a new subtype of hubs not previously described. Proteins with multiple interaction surfaces have previously been recognised as a distinct type of hub that can interact with multiple partners simultaneously. However in our pathway network we find hubs that can be composed of clustered proteins each with multiple interaction surfaces that can interact with multiple proteins simultaneously. There is also another possible type of clustered-hub composed of proteins with one interaction surface that might be able to interact with multiple proteins, but only at different times or locations. This second type of protein is likely to contain an interaction domain like an SH3 domain, SH2 domain, EH domain, PTB domain or adaptor appendage domain, that can bind to short sequence motifs dispersed in the interaction partners. From a pathway-centric viewpoint we do not consider proteins with sequential interactions as functional hubs unless they are clustered.
|
|
|
Design principles and spatio-temporal dynamics behind pathways that can aid in understanding pathways and in experimental design and interpretation |
1. Predicting the timescale of events in a pathway from its interactome plot
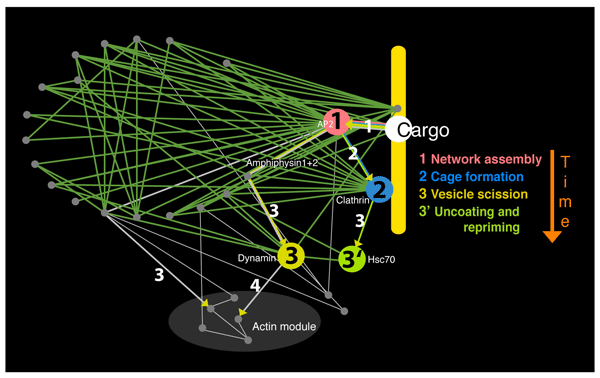 |
- A short path-length gives an immediate response
- To put a time delay in the response an additional interaction step is added: thus an extra path-length/link gives a molecular-clock delay to this event
|
How the Time-line works:
Starting with the initiation point of CME (which is cargo binding) we count the number of links required to get from one protein of interest to another. In this analysis protein complexes such as AP2 are treated as a single node and this type of analysis discounts the effects of protein concentration and affinities and so is an approximation.
We can give the approximate time line of events in CME by concentrating on path-lengths in the pathway protein interactome. There is one path-length between cargo and AP2, there are two path-lengths connecting cargo to accessory proteins and clathrin, and there are three path-lengths to dynamin and Hsc70. Dynamin is connected into the network through accessory proteins such as snx9, intersectin and amphiphysin (amphiphysin shown in figure above) and not via the clathrin hub, as dynamin function is spatially separated from clathrin (dynamin acts at the neck of the nascent vesicle). In vivo fluorescence studies of clathrin coated pit dynamics validate the time line (see below).
In vivo fluorescence studies of clathrin coated pit dynamics validate the time line where dynamin and auxilin/Hsc70 are seen to act just before clathrin spots leave the visualisation field in total internal reflection microscopy experiments. The actin machinery comes into play after this, where actin modulators in our pathway protein interactome would have a path-length of three to four from cargo binding. While overexpression of labelled proteins can give much information about the pathway, there are limitations of these visualisation approaches. Here we can be informed by the pathway protein interactome. Firstly, the pathway protein interactome time line gives information on early events that are difficult to probe with fluorescent markers. Clathrin is generally the marker used to identify coated pit formation, and so events occurring before clathrin begins to polymerise may well be difficult to identify, since in early stages of network assembly AP2 may not be as highly concentrated and may be much more dynamic. Even after clathrin is detected as a spot the ability to detect AP2 in this spot will depend on the labelling efficiency. Secondly, the pathway protein interactome tells us that overexpression of nodes will frequently cause the system to work sub-optimally if is able to disrupt a hub and so the resulting data should be treated with care.
|
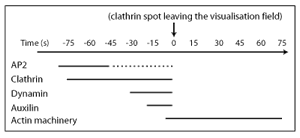
These data come from in vivo imaging using total internal reflection fluorescence microscopy from the following papers: (AP2, clathrin: Ehrlich et al 2004, Cell 118; Rappaport et al 2006, Am J Physiol 291) (dynamin, clathrin: Merrifield et al 2002, Nat Cell Biol 4; Soulet et al 2005, Mol Biol Cell 16) (auxilin, clathrin: Massol et al, 2006 PNAS 103; Lee et al 2006 J. Cell Sci, 119) (actin, clathrin: Merrifield et al 2005, Cell 121; Yarar et al 2005, Mol Biol Cell, 16). |
|
2. Some implications for experimental design:
In the thread and nail picture to the right, removal of key nails (hubs) would cause the picture to unravel, but deletion of minor nails (accessory nodes) will have little consequence. In interactome it has already been shown that the most connected proteins are likely to be the most essential. Our analysis makes a number of refinements to this picture.
- the most connected proteins (hubs in an interactome) are not necessarily hubs in a the cell. For example, dynamin binds to many partners in different contexts in the cell and so should be represented in the interactome as multiple copies and should not be a hub-point in the network. Currently we do not know how widespread this observation may apply.
- overexpression of any individual accessory node that links to a hub in an interactome would automatically lead to a reduced incorporation of other accessory nodes and so can be a more positive test for the role of a protein in a pathway (see powerpoint presentation below for more information on this concept, and supplementary material of Schmid and McMahon (Nature 2007) for detailed description and implications for experimental design).
|
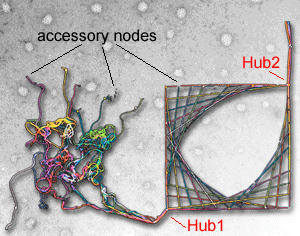
Thread and nail picture by Jessica Thurlbourn superimposed on an electron micrograph of clathrin-coated pits |
3. A new mode of interaction: Matricity
Directionality can be driven through changing interaction modes
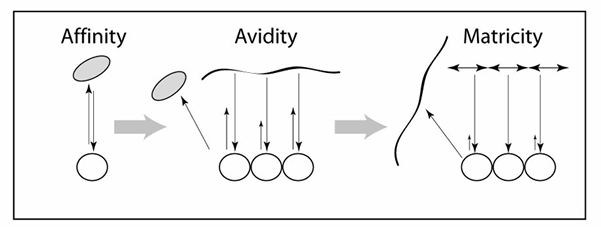 |
Affinity-driven interactions have equal on- and off-rates whereas in avidity-driven interactions the off-rates are significantly reduced due to multiple interaction points. The third form of interactions, matricity-driven interactions, involve a rigid matrix which leads to a further reduction in off-rates or the absence of off-rates altogether (in our case polymerised clathrin has much less flexibility than any of the accessory proteins). These interactions are illustrated in the powerpoint presentation below and more extensively in our paper (see pathway-progression in Schmid and McMahon Nature 2007).
|
4. Inherent Flexibility in pathways:
A hub-centric pathway has the advantage of easily being able to add additional modules to the system (see Schmid and McMahon 2007, for adding modular attachments or 'plug-ins'). In CME, introducing alternative cargo adaptors is such an example, where a different cargo can be internalised using the pathway by simply adding on another accessory node to the central hub that can work as a cargo adaptors. Moreover, addition of such alternative adaptors could also be an evolutionary consideration, to ensure that the retrieval of some ligands is less dependent on adaptors like AP2 which could well be partially occupied by other activated receptors. It has been noted previously by network biologists that alternative routes provide robustness and account for apparent redundancy in networks.
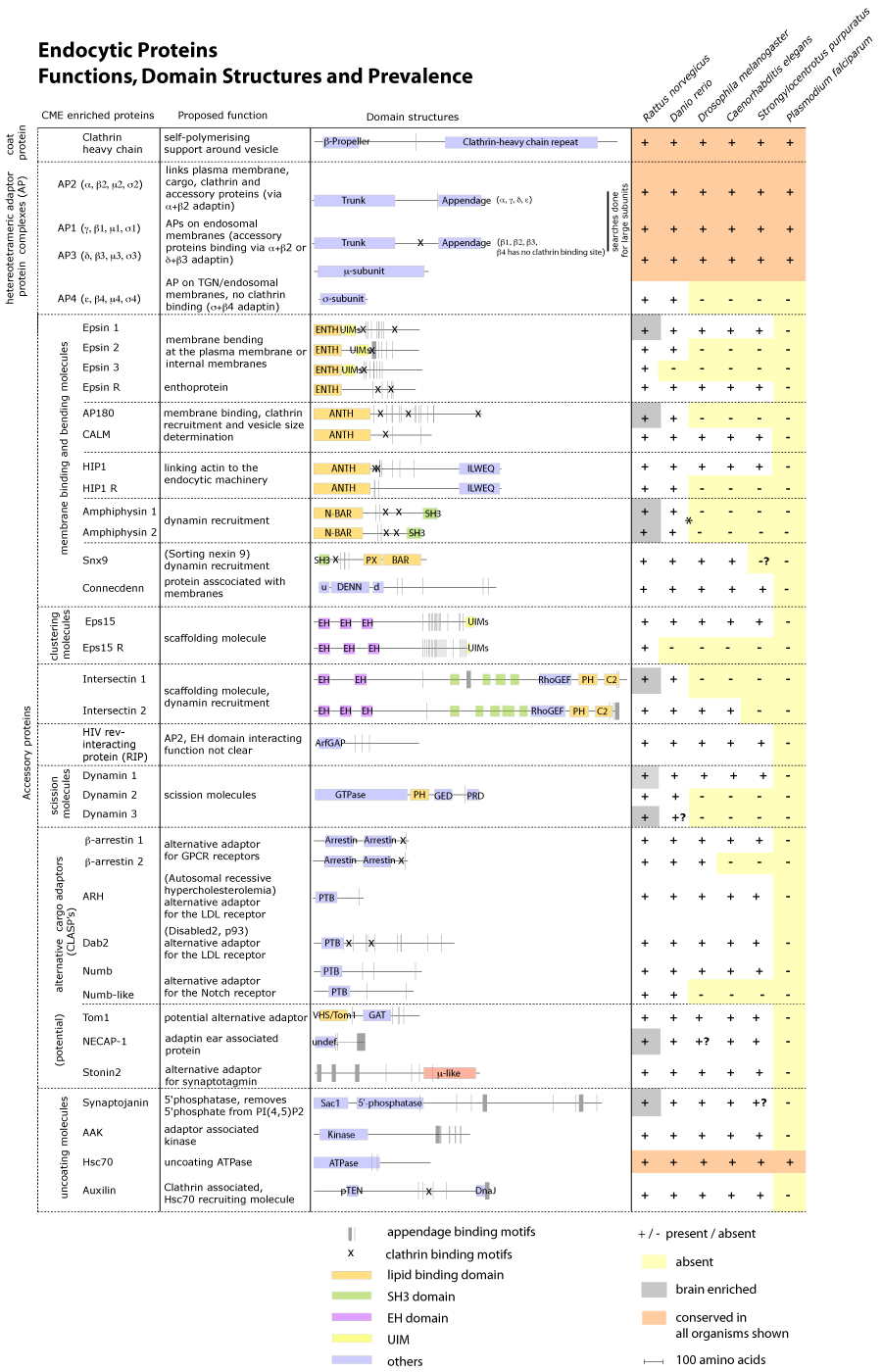
We investigated the conservation of the CME network across species in great detail and observed that conservation of endocytic proteins through the animal kingdom was high and the connectivity of accessory proteins is also mostly conserved (see Table below). In lower organisms there is less duplication of proteins and thus less redundancy and so knock-out phenotypes tend to be stronger. There is a trend for higher eukaryotes to have brain-specialised isoforms as well as ubiquitous forms of many proteins, while one form of the protein seems to suffice in other multicellular organisms with nervous systems such as Drosophila melanogaster, Caenorhabtidis elegans, and Strongylocentrotus purpuratus (sea urchin). It is tempting to suggest that these specialised forms of many node proteins may have allowed the development of the brain as we know it, and would appear to have arisen from gene duplication events in higher organisms. It is interesting that there is also a duplication of genes for exocytic components and synaptic vesicle proteins (side observation). It would be interesting to know if these specialised proteins form a transcriptional cluster to make it easier to turn on all these proteins in the brain.
|
 |
|
Our publications on this subject
This page was written to accompany an Analysis paper in Nature 2007, 448, 883-888 (see abstract)
but Praefcke et al EMBO J. 2004; Schmid et al PLoS Biol. 2006; Schmid and McMahon Nature 2007 are all relevant. |
